SISTEMA INNATO COMO DEFENSA DE AGENTES INFECCIOSOS
 Un individuo está expuesto a
muchos agentes infecciosos; sin embargo, la mayoría de los microorganismos
infecciosos no logran ingresar al individuo, gracias a las barreras físicas y
químicas que éste presenta. La barrera física más importante es la piel; la
integridad de ésta, junto a la secreción de mediadores químicos, evita el
ingreso de microorganismos patógenos. Asimismo, las mucosas poseen una serie de
atributos (secreciones, flujo ciliar) que dificultan el ingreso de
microorganismos por esa vía. Además, la existencia de poblaciones microbianas
no patógenas residentes (flora normal), también impide la colonización de las
mucosas por agentes infecciosos.
Un individuo está expuesto a
muchos agentes infecciosos; sin embargo, la mayoría de los microorganismos
infecciosos no logran ingresar al individuo, gracias a las barreras físicas y
químicas que éste presenta. La barrera física más importante es la piel; la
integridad de ésta, junto a la secreción de mediadores químicos, evita el
ingreso de microorganismos patógenos. Asimismo, las mucosas poseen una serie de
atributos (secreciones, flujo ciliar) que dificultan el ingreso de
microorganismos por esa vía. Además, la existencia de poblaciones microbianas
no patógenas residentes (flora normal), también impide la colonización de las
mucosas por agentes infecciosos. 
INMUNIDAD INNATA
La respuesta inmune innata brinda la primer línea de defensa contra los microorganismos
invasores pero además provee el contexto biológico y las señales que instruirán al sistema
inmune adaptativo para montar su respuesta. Así, los eventos ocurridos en el contexto de la
inmunidad innata, determinan el perfil del tipo de respuesta adaptativa que se desarrollará
contra el agente patógeno.
El objetivo de la inmunidad innata es evitar la instalación del proceso infeccioso; si éste se
produce, dicho mecanismo inmunitario logra establecer un ambiente para que se desarrolle
una respuesta adaptativa. Los mecanismos efectores de defensa de la inmunidad innata están
compuestos por células que cumplen funciones defensivas (fagocitosis, citotoxicidad) y factores
solubles (citoquinas y quemoquinas, interferones, complemento) que controlan y destruyen
los microorganismos que ingresan.

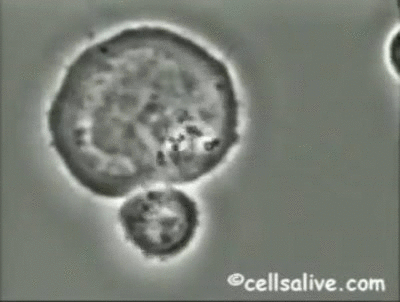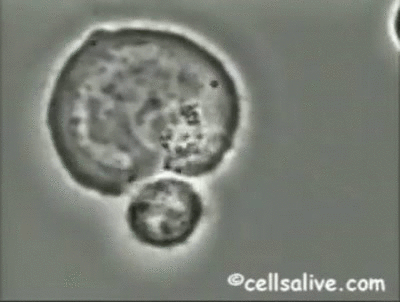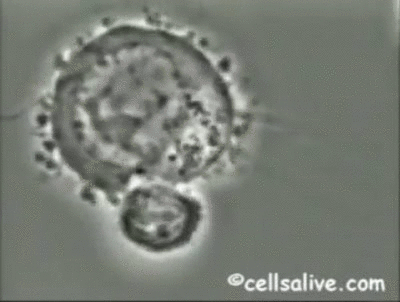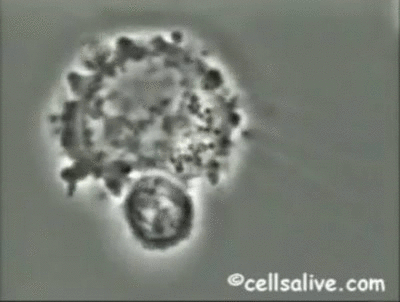
Los mecanismos fundamentales de la inmunidad innata operantes contra bacterias extracelulares
son la fagocitosis, la respuesta inflamatoria y la activación del complemento.
Los fagocitos pueden unirse a bacterias extracelulares mediante una serie de receptores;
dicha interacción, junto con la señalización intracelular realizada por los TLRs, activa los
fagocitos incrementando su capacidad fagocítica y microbicida. De ahí que la resistencia de
las bacterias a la fagocitosis y a la digestión dentro de los macrófagos, es un determinante
importante de la patogenicidad y virulencia de las mismas.
La activación de los fagocitos
también provoca la secreción de citoquinas proinflamatorias como el factor de necrosis tumoral
(TNF-a) y las interleuquinas IL-1, IL-6 e IL-8, que inducen la adhesión de neutrófilos
y monocitos al endotelio vascular en el sitio de la infección, seguida por la migración, acumulación
local y activación de las células inflamatorias que eliminan las bacterias. Sin embargo, la producción de grandes cantidades de citoquinas puede ser perjudicial y
de hecho son responsables de algunas manifestaciones clínicas de las infecciones por bacterias
extracelulares. El daño de tejidos normales adyacentes es un efecto colateral de estos
mecanismos de defensa. La consecuencia más grave inducida por la secreción descontrolada
de citoquinas, es el shock séptico que puede presentarse con coagulación intravascular diseminada,
falla multiorgánica y muerte, propio de algunas infecciones por bacterias gramnegativas
(desencadenado por el LPS) y grampositivas (donde el peptidoglicano y los ácidos teicoicos
desencadenan efectos similares).
La activación del complemento en ausencia de anticuerpos, también tiene un rol importante
en la eliminación de estas bacterias. El peptidoglicano de las paredes celulares de
las bacterias grampositivas y el LPS de las paredes celulares gramnegativas, activan la vía alterna del complemento promoviendo la formación de C3 convertasa.

INMUNIDAD FRENTE A BACTERIAS INTRACELULARES
INMUNIDAD INNATA FRENTE A LOS VIRUS
Los virus son microorganismos intracelulares obligados, que generalmente ingresan a las células susceptibles usando como receptores las moléculas normales de superficie celular. Cuando el virus está dentro de la célula huésped, causa lesión celular por diferentes mecanismos. La replicación viral puede interferir con la síntesis proteica celular y provocar la muerte de la célula por lisis, liberándose muchas partículas virales nuevas (virus citolíticos).
La inmunidad contra los virus debe ser capaz de actuar en las distintas poblaciones de células infectadas (dado que distintos virus infectan distintas células). Dicho mecanismo inmunitario opera a dos niveles: previo a la invasión celular, en la etapa inicial de la infección y después de la invasión cuando los virus son inaccesibles a los anticuerpos y fagocitos.

En primer lugar, la infección viral estimula la producción, por parte de las células infectadas, de IFN tipo 1 (que comprende dos grupos serológicamente distintos, el α y el β). El IFN tipo 1 tiene muchas acciones biológicas. En principio inhibe la replicación viral estimulando la síntesis de enzimas celulares que interfieren con la replicación del ácido ribonucleico (ARN) o desoxirribonucleico (ADN) viral.
Su acción antiviral también es ejercida sobre las células vecinas no infectadas, que quedan así protegidas de la infección. Además, el INF tipo 1 inhibe la proliferación celular por inducción de las mismas enzimas mencionadas anteriormente y de otras que actúan inhibiendo la síntesis de aminoácidos. También aumenta el potencial lítico de las células NK cuya función principal es matar las células infectadas por virus. Por último, modula la expresión de moléculas MHC, aumentando la expresión de las moléculas MHC clase I e inhibiendo las de clase II. Así mejora la eficiencia de los linfocitos T citolíticos que reconocen antígenos extraños asociados a moléculas MHC de clase I.
En segundo lugar, las células NK lisan muchas células infectadas por virus, constituyendo uno de los mecanismos efectores principales en los estadíos iniciales de la infección viral.Además del IFN tipo 1, el IFN-γ, el TNF y la IL-2, aumentan el potencial lítico de estas células.

BIBLIOGRAFÍA
http://www.tirsoferrol.org/ciencias/pdf/a21_inmunologia.pdf
http://www.higiene.edu.uy/cefa/2008/Inmunidadcontralosagentesinfecciosos.pdf
No hay comentarios:
Publicar un comentario